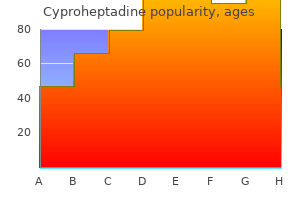
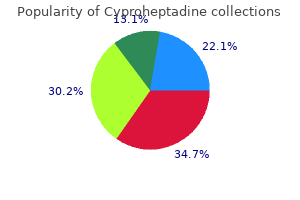
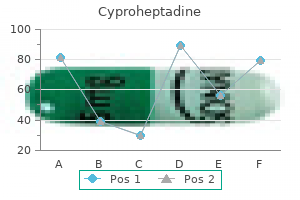
Only recently have we been able to test these ideas in nature and say with confidence how real species in nature have formed allergy testing ashby de la zouch cheap 4 mg cyproheptadine. Since Darwin the role of natural selection has been of great interest allergy symptoms but not allergic to anything generic cyproheptadine 4mg otc, and we know more about its role than that of any other process allergy kiwi buy generic cyproheptadine 4 mg on line. Such isolation can occur when separate populations adapt to different environments (ecological speciation) as different alleles favored in one environment but not the other gradually accumulate between populations allergy high buy discount cyproheptadine 4mg. Alternatively allergy symptoms body aches 4mg cyproheptadine sale, selection may build genetic differences among populations that experience similar selection pressures if the populations by chance experience and accumulate different sets of advantageous mutations (mutation-order speciation) allergy report oklahoma cheap 4 mg cyproheptadine with visa. Biotic interactions with other species allergy symptoms of the throat buy cyproheptadine 4mg, including predatorprey allergy shots denver purchase 4 mg cyproheptadine fast delivery, host-parasite, competition and mutualism, are also a major (perhaps the major) source of selection on populations. This process, called reinforcement, represents the only known circumstance in which natural selection directly favors the evolution of stronger reproductive isolation. Otherwise, as described earlier, the role of selection is indirect- reproductive isolation evolves as an incidental consequence of adaptation. Since the exaggeration of such traits is caused by sexual selection, it seems likely that sexual selection is also frequently involved in speciation. Any such role would likely involve natural selection, too, because it is the process that leads to divergence of mate preferences or that favors the evolution of traits that ameliorate intergenomic conflict. Sometimes, mutations of large effects on reproductive isolation are found, whereas reproductive isolation often results from the accumulated effects of many small-effect mutations. Genetic "signatures" of selection detected on speciation genes provide some of the best evidence that natural and/or sexual selection have been responsible for driving the mutations to high frequency, and hence for the evolution of reproductive isolation. We are beginning to learn why speciation genes are often clustered rather than dispersed within the 486 Speciation and Macroevolution exploit them. A surprisingly common mechanism of sudden speciation is via the evolution of polyploidy. The process is most common in plants, but several examples from animals have recently been discovered. Classic examples include the finch radiations on the Galapagos and Haґ waiian Islands. In both cases the species have evolved a wide diversity of beak sizes and shapes that enhance the ability of individuals to exploit particular resources, such as hard seeds, nectar from long-tubed flowers, or insects under bark. In the few adaptive radiations that have been studied intensively, it is clear that the same natural selection pressures that adapt populations to distinct niches also indirectly contribute to the buildup of reproductive isolation between populations. In these few studied cases, at least, there is a close connection between rapid speciation and adaptive evolution. Adaptive radiations are particularly prevalent where ample resources are available, and few competing lineages take full advantage of them (ecological opportunity). Even under such conditions, however, some lineages diversify more readily than others, as though they have intrinsic differences that affect their abilities to speciate rapidly, or to adapt to and usurp, novel resources. For example, it has been proposed that the huge diversity of angiosperm plants is attributable to the evolution of the flower. Adaptation of flower structures to different suites of pollinators in different environments might speed the evolution of premating reproductive isolation. Another hypothesis is that the evolution of traits permitting certain insects to consume plant tissue is behind the astonishing diversity of phytophagous insects, such as herbivorous beetles, found today. Plants are incredibly abundant and diverse in their leaf structures, chemistries, and life histories, which favors niche specialization and diversification by insects that Adaptive radiations represent episodes of particularly fast evolution and speciation. In contrast, study of patterns of evolution in the fossil record and in phylogenetic trees has found that evolution is often slow. Lineages frequently undergo long periods in which little evolution seems to take place-at least in easily identified morphological traits (perhaps rates are not so slow in other aspects, such as at genes involved in fighting disease). The hypothesis of punctuated equilibria was an extreme statement about rates of evolution in nature: that evolution hardly ever occurs except in the relatively brief periods during which speciation also takes place. The rest of the time, so the hypothesis goes, species exhibit stasis, changing little. A key question is whether the punctuated equilibrium is a caricature of evolutionary patterns in the fossil record. Sustained directional changes in traits might indeed occur infrequently and episodically, but the rest of the time evolution might be better described as oscillating rather than static, or at least not sustained and directional, with fluctuations of varying amplitude taking place through time. If a relationship between a trait possessed by species and speciation or extinction rates holds consistently across multiple lineages and over time, the result will be a large-scale increase in the prevalence of that particular trait in nature. We can think of this process as species selection-the macroevolutionary analogous of natural selection on individuals within populations. The notion of species selection has been controversial, and many researchers regard it as a weak force compared with ordinary natural selection within species. However, species selection need not oppose ordinary natural selection, and it may indeed generate trends in the absence of any net direction to evolution within species. Dinosaurs possessed exquisite adaptations to the many environments in which they occurred, yet they were wiped out en masse by catastrophic environmental changes never before experienced during their many millions of years of history. Indeed, it does not often happen that the lineages that dominatedthe earthprior to mass extinctions recover and reassume their dominant positions afterward. The macroevolutionary processes of speciation and extinction, of species selection and species drift, acting over long spans of time created the biodiversity that has assembled into ecological communities and continues to influence how local assemblages change through time. Natural selection on variation within species has produced the myriad adaptations of species to one another and to the abiotic environments they encounter across their geographic ranges. As species adapt to one another, the strength of their interactions changes and the flow of energy and materials is altered, producing consequent changes in the properties and dynamics of the surrounding ecosystem. Thus, microevolution on species within communities generates new species and modifies the traits that species possess and so provides the material that drives macroevolutionary changes. Studying speciation Species are the fundamental units of biodiversity, but the definition of a species remains a subject of debate within evolutionary biology. One resolution of this debate views alternative species definitions as different stages in the process of speciation, in which conspecific populations diverge, accumulate intrinsic barriers to gene exchange, and ultimately become exclusive or reciprocally monophyletic groups. Most studies of speciation have focused on the evolution of reproductive isolation or intrinsic barriers to gene exchange. Such barriers may result from a variety of trait differences, some of which are simply a by-product of divergence in allopatry. Barriers may prevent individuals from meeting or mating, they may compromise gamete interactions, or they may reduce the viability and/or fertility of hybrid offspring. Occupying different geographic regions; geo- species are widely viewed as fundamental units of biological organization. However, the defining qualities of species, the nature of the boundary between species, and even the reality of species remain matters of dispute. It is ironic that the concept or definition of species, so central to the studies of evolution, ecology, and conservation biology, has engendered so much confusion and debate. In contrast, the rules for naming species (for animals embodied in the International Code of Zoological Nomenclature) are very clearly described and widely accepted. The last two decades have witnessed gradual acceptance of the view that there is not one "right" species concept or definition. One approach to resolving past disagreements is to recognize the difference between a species concept and a species definition. The former is what is meant by the word species; the latter involves defining the criteria used to delimit species. Because species are defined over time as well as space, and because speciation is a process not an event, differences among species definitions may then reflect different landmarks along the path from conspecific populations to separate species. Movement or incorporation of alleles from one population into one or more different populations. A group of organisms (taxa) that all share a most recent common ancestor not shared by any other organisms (taxa). The (usually diploid) cell formed by the union of two (usually haploid) gametes. Like cells or individual organisms, the evolutionary biology literature presents a bewildering array of different species concepts or definitions (as many as 24 have been identified). Some of the concepts are subtle variations on basic themes, but at least seven major concepts can be differentiated along a number of axes, including whether they are retrospective (species as products of history) or prospective (species as lineages extending into the future), whether they are relational or nonrelational, whether they are based on pattern or process, and the extent to which they are operational, 490 Speciation and Macroevolution Table 1. Major species concepts or definitions Biological Species Concept "Groups of actually or potentially interbreeding natural populations which are reproductively isolated from other such groups. Character-Based Concepts Perhaps the simplest and most intuitive concepts of species are those that are "character based. Of course, care must be taken to ensure that phenotypically distinct groups do not simply represent differences between males and females or differences among life history stages. Although easy to apply, character-based definitions view speciation as equivalent to divergence. If isolated populations diverged as a result of natural selection that led to local adaptation, or if allele frequencies drifted to fixation for different alleles at a gene locus, these populations would be viewed as distinct species. Strict ap- plication of such character-based concepts might result in a tremendous proliferation of species and the elevation of many current subspecies or races to species status. Darwin clearly recognized that evolution (including the origin of species) is a process and that sampling diversity at any one point in time should reveal populations at all stages in the process. Indeed, Darwin saw the continuum of differences between populations as direct evidence for the evolutionary process. He wrote: Certainly no clear line of demarcation has as yet been drawn between species and sub-species. These differences blend into each other in an insensible series; and a series impresses the mind with the idea of an actual passage. Thus, species boundaries may be fuzzy and develop gradually, with few rules about how much difference needs to accumulate before lineages should be recognized as different species. However, for allopatric populations, reliance on amount (or quality) of difference seems to be the only possibility. Darwin commented that in cases "in which intermediate links [between populations] have Species and Speciation not been found. This genotypic cluster definition is also character based, but by examining character differences in sympatry, it attempts to assess whether the existing differences persist. The Biological Species Concept (or Isolation Concept) 491 flow of alleles between the two populations/species. The bright side is that observed patterns of differential gene flow (introgression) can be used to gain insight into the genetic architecture of speciation. Species are characterized by interbreeding within a population and reproductive isolation among groups of populations. Reproductive isolation is a consequence of intrinsic barriers to gene exchange, barriers due to the properties of the organisms themselves and not simply geographic separation. The presence of intrinsic barriers to gene exchange suggests that two species will persist, although ecological differentiation may also be a prerequisite for coexistence. First, it is difficult to estimate amount of gene flow in natural populations; therefore, absence of gene flow is often inferred from patterns of differentiation for genotypic or phenotypic markers. Patterns of gene exchange vary not only in space and time but also across the genome. In some regions of the genome there may be no gene exchange, whereas in other regions, hybridization results in the Descent with modification produces a nested hierarchy of traits and taxa. The structure of this nested hierarchy can be revealed by phylogenetic analysis, which documents the pattern of branching events (forward in time) or coalescent events (backward in time) that define extant individuals, populations, or species. A phylogenetic or genealogical perspective suggests that species should be considered to be monophyletic or exclusive groups. An exclusive group is one whose members are all more closely related to one another than to any individual outside the group. The genealogical species concept of Baum and Shaw argues that exclusivity is an important criterion for species status. It seems quite reasonable that all members of a species should be closely related, but because of ancestral polymorphism and incomplete lineage sorting and/or ongoing hybridization, individuals within a species may, in fact, be more closely related to individuals of other species. Relationships depend on which gene or gene region is sampled, and exclusivity may characterize some genome regions and not others. It is not clear what proportion of the genome must be "exclusive" before a group is considered a genealogical species. According to a strict definition, many entities now viewed as independent evolutionary lineages would be considered conspecific, because genealogical speciation requires very long periods of time. Evolutionary Species Concept For most of evolutionary history, the only data we have about species and speciation come from the fossil record. Thus, the data are purely phenotypic (usually morphological), and neither the nature nor the quantity of the data allow direct assessment of gene exchange or exclusivity. A very different and more general view that has been applied to fossil data is the evolutionary species concept, originating with the paleontologist G. This concept defines species as populations through time (which can be followed in the fossil record) that maintain their separate identity (in the presence of other lineages) and exhibit an independent evolutionary trajectory. Eventually, some of the fixed differences affect the ecology, behavior, physiology, or reproductive biology of the diverging lineages. As a by-product of this divergence, the populations accumulate differences that affect the probability of their interbreeding or the success of their progeny should an individual mate with a member of the "other" population. However, these populations are exclusive groups only at some (perhaps relatively few) regions of the genome. Over time, an increasing proportion of the genome diverges, and each of the diverging populations becomes an exclusive group across the genome. The critical event in this life history is the evolution of reproductive isolation, the appearance of intrinsic barriers to gene exchange. This transition alters the outcome if or when secondary contact occurs between the diverging populations. Should the populations become sympatric, intrinsic barriers will prevent gene flow and the erosion of genetic differences. Similarly, in models of sympatric speciation, the evolution of reproductive barriers leads to the cessation of gene flow and enables further divergence of two subpopulations. Evolutionary biologists who study speciation therefore focus on the nature of intrinsic barriers to gene exchange: what they are, when they act in the life cycle of the organism, to what extent they reduce gene flow, and when in the history of divergence they arose. Answers to these questions emerge from comparisons of closely related (recently diverged) species and "incipient" species. Fixed differences, reproductive isolation, exclusivity, and separate identities are all patterns or characteristics that must be defined in terms of differences between individuals and populations. A number of evolutionary biologists have argued that species concepts should be nonrelational, defined in terms of what is shared, rather than what is different. The recognition concept defines species as populations of sexual organisms that share a common fertilization system or specific mate recognition system. The emphasis is on defining an interbreeding unit, a "field for recombination," a group of organisms held together by "genetic cohesion. In practice, defining cohesion mechanisms is virtually equivalent to defining reproductive isolation. In either case, a group of individuals has to be partitioned into two or more subgroups, each of which shares fertilization and mate recognition systems within the subgroup but differs in these respects from other subgroups. But if two species are to persist in sympatry, the competitive exclusion principle from ecology says that they must be ecologically distinct. One exception is the cohesion species concept, which defines species as "the most inclusive population of individuals having the potential for phenotypic cohesion," which is mediated by both genetic and demographic exchangeability. Groups of organisms that are demographically exchangeable are ecological equivalents. This concept is of particular use in sympatric asexual lineages, in which even in the absence of any gene exchange, demographic exchangeability implies that lineages belong to the same species. It is useful to think of the process of divergence as the life history of a species. Fixation of alternative alleles or phenotypes in the two populations results in their being considered Many phenotypic differences between diverging lineages can result in barriers to gene exchange. The traditional approach to classifying such barriers is to organize them with respect to whether they act before mating and/ or zygote formation or whether they are a consequence of the reduced fitness (viability, fertility) of hybrid offspring. Most classifications recognize three distinct sorts of barriers: (1) premating, (2) postmating but prezygotic, and (3) postzygotic. Premating Barriers Premating barriers are those that result from trait differences that prevent hybridization between distinct species. Species and Speciation Barriers to gene exchange will exist if potential mates do not meet (temporal and habitat or ecogeographic isolation), if potential mates meet but do not mate (behavioral isolation), or if attempted copulation does not result in sperm transfer (mechanical isolation). Premating barriers to gene exchange have been studied in many different animal and plant systems, revealing a host of different mechanisms whereby gene exchange is limited or prevented. Temporal isolation reflects seasonal or diurnal differences in the times at which adults are present or sexually active. Thus, flowering time differences in plants, and major life cycle differences in animals. In many marine invertebrates, mating (in the narrow sense) does not occur, and eggs and sperm are simply broadcast in the water column. Spawning times can be determined by lunar cycles, so that in some corals, eggs and sperm from closely related species are unlikely to encounter each other in the water column. Similar patterns are seen in some moths, in which sexual activity is limited to a relatively narrow window in the diurnal cycle and may be displaced from the corresponding window for a sympatric close relative. Habitat or resource isolation results from the association of particular populations or lineages with specific habitats or resources. Observed associations can be the result of differential adaptation to habitats or differential preference for habitats. Many examples come from the insecthost plant literature, in which insect lineages have apparently diversified by adapting to new host plants, resulting in reproductive isolation between the derived forms. There are also numerous examples of plants with different habitat needs or requirements; some of the best studied involve adaptation to different soils. Geographic isolation can result from local adaptation if habitats or resources are geographically separate; this phenomenon has been termed "ecogeographic isolation. If ecological factors do not prevent individuals from meeting, then behavioral differences may well prevent them from mating. Many examples have been documented of species-specific communication systems that function in mate finding and mate recognition.
Surely adaptation occurred allergy symptoms 5 months generic cyproheptadine 4 mg with amex, but Kimura argued that it occurred in only a small proportion of the genome at any one time and that natural selection was unlikely to account for the maintenance of extensive molecular variation observed within species and for the fixation of variation between species allergy forecast lancaster pa generic cyproheptadine 4mg line. It was also inferred that many mutations were in fact harmful and eliminated from populations allergy forecast today generic cyproheptadine 4mg without a prescription, so those regions of genes and the genome that were functionally critical would remain largely invariant allergy medicine vertigo discount cyproheptadine 4mg. If selection could not reasonably explain these observations allergy medicine poison ivy buy cyproheptadine 4 mg without a prescription, 370 Genes allergy symptoms from pollen purchase 4 mg cyproheptadine amex, Genomes allergy shots vs antihistamines purchase 4mg cyproheptadine otc, Phenotypes rate of evolution allergy medicine and grapefruit juice discount cyproheptadine 4mg with mastercard. For new neutral mutations destined to be fixed by drift, average time to fixation (in units of generations) is approximately four times the long-term population size. Thus, for large populations, we expect long times to fixation, and thus lots of "transient" genetic variation in populations drifting slowly through them. Together, these processes mean that the level of variation within species is a function of population size and mutation rate. For a stable population, the balance of new mutations and loss or fixation by drift leads to an equilibrium level of variation. It can be shown that this level of variation is such that the probability that an average nucleotide site shows a difference between two randomly chosen chromosomes (or is heterozygous in a randomly chosen diploid sexually reproducing organism) is approximately equal to four times the long-term population size multiplied by the rate of mutation. The amount of divergence between two sequences sampled from two different species will be equal to the mutation rate times twice the time since speciation plus an additional amount equal to the expected number of differences between two randomly chosen chromosomes in the ancestral population. Because variation between species is but an extension of variation within species, and both are ultimately driven by mutation, then strictly neutral variation within species should be positively correlated with strictly neutral variation between species. This broad concept became known as the neutral allele theory of molecular evolution, and it has formed a conceptual and model framework on which much of the current field has been based. Perhaps most important has been the recognition that all populations of organisms are finite in size, so that the stochastic process of genetic drift forms a background on which all other evolutionary forces act. The term neutral theory (as it is often called) can be misleading, as not all variation is selectively neutral; rather, the theory allows that a significant portion of new mutations are strongly deleterious and nearly immediately removed from the population. And the theory does allow for a limited number of adaptive mutations; however, the remaining mutations are selectively equivalent (neutral), and their dynamics are determined solely by genetic drift. Thus, the majority of variation we see within and between species is assumed to have no effect on fitness of organisms. The availability of these correlated evolutionary histories allowed for the development of new statistical and computational approaches for testing models of molecular evolution, particularly the neutral theory. The field was no longer theory rich and data poor, as now the data began pouring in at an astounding rate. The strict neutral theory was clearly an oversimplification but has provided the field with a valuable reminder of the importance of stochastic processes in all populations and a valuable null hypothesis against which to evaluate data. And the rate at which new alleles become fixed in a population (the substitution rate) is essentially equal to the "neutral" mutation rate per generation. Thus, if the neutral mutation rate remains constant, so should the Mutations that confer a fitness advantage will increase in frequency in the population because of positive selection. If the variant goes to a frequency of 100 percent, the population has now undergone a substitution of one variant for another. Positive selection can lead to very rapid rates of fixation, orders of magnitude faster than the rate of fixation due to genetic drift alone. Because adjacent nucleotides are tightly linked genetically, selection impacts not only the beneficial mutation but also the region of the genome in which that variant is located. Rapid fixation can therefore fix not only the favored variant but also the surrounding segment of the genome, resulting in a "selective sweep" and consequently a genomic region of initially no or very reduced adjacent neutral variation. Perhaps surprisingly, the average divergence of linked neutral sites is unchanged from the neutral prediction. Such patterns provide much insight into the frequency Molecular Evolution and location of adaptive fixations throughout the genome (see chapter V. While the rate of new mutations in the population is unchanged with natural selection, the fixation rate for beneficial mutations is dramatically increased, leading to increased sequence divergence between species for those specific sites under positive selection compared with adjacent neutral variants whose dynamics are determined by stochastic processes of genetic drift alone. The study of protein-coding sequences provides a particularly illustrative, and useful, example of how this contrasting pattern of positively selected, negatively selected, and neutral variation can be used to infer where and how natural selection has acted in the genome. While there are 61 possible combinations of three nucleotides that encode amino acids (three additional encode protein synthesis "stop" signals), there are only 20 common amino acids. Those triplet codons that encode the same amino acid differ by what is known as synonymous or silent variants. Those that result in a change in the encoded amino acid are termed nonsynonymous or replacement variants. Since protein function is largely determined by its amino acid sequence, the fitness consequences of nonsynonymous mutations are much greater than those of synonymous mutations. Constraints on protein function result in strong purifying selection on nonsynonymous variants, preventing them from reaching substantial frequencies in populations (polymorphism) or from going to fixation (substitutions). Some proteins play key roles in adaptation to new enzyme substrates or respond to biotic or abiotic challenges. Here, positive selection favoring new amino acid variants leads to the accelerated fixation of mutations. Some adaptive responses require repeated changes in protein sequence (such as at the antigen-binding sites of some immunity proteins), resulting in successive accelerated replacements. Somewhat counterintuitively, such positive and negative selection has little to no effect on the rates of substitution of adjacent strictly neutral variants. Thus, contrasting levels of variation and/or divergence at nonsynonymous to synonymous sites can provide estimates of the strength of both positive and purifying selection for a proteincoding gene. The ideal neutral benchmark is found in "dead genes" that no longer function (pseudogenes), in which levels of variation and divergence are usually close 371 to those seen at synonymous sites. These contrasts of nonsynonymous and synonymous variation and divergence form the basis of several tests to detect natural selection acting on genomes and uncover the functional targets of that selection (see chapter V. Not all mutations are simply strictly neutral, lethal, or strongly favored; rather, functional and population genetics studies have demonstrated that many mutations affect function only slightly, most in a slightly negative manner but sometimes improving function a bit. In these instances, whether a mutant acts as a neutral variant can be influenced by the population size. Consideration of the relative strength of natural selection and genetic drift reveals that if the difference in reproductive success (fitness) is less than the reciprocal of the long-term population size, then the mutant will behave as a neutral variant, even if it would have a (slight) selective advantage or disadvantage in an infinitely large population. One impact of this class of variants is that fluctuations in population size among lineages, or even along lineages, leads to fluctuations in the ability of natural selection to "see" these variants. This phenomenon is most clearly observed in the "generation time effect" observed in the molecular clock for some types of variants. As predicted then, rates of protein evolution are slower on a per generation basis in short-generation mammals than rates of substitution in long-generation mammals. For example, these valuable studies have provided real data sets with known phylogenies with which to evaluate the accuracy of statistical methods to estimate the phylogenetic relationships of organisms and their ancestors when we have only sequence data from the extant end points of the evolutionary process for study (figure 2). Experimental microbial and viral evolution studies are also allowing direct tests of evolutionary hypotheses regarding adaptation, including 372 7 Genes, Genomes, Phenotypes R 1 6 6 9 6 3 4 5 Actual 3 J 9 0 M K 6 3 1 L N Q P O 6 R 1 3 Estimated J 0 10 6 5 7 78 1 23 M 6 3 1 K L N 67 Q P O Figure 2. Mutational change in virus cultures demonstrates descent with modification and molecular evolution, and the molecular clock concept. This population was initiated with one virus and split in binary fashion into several sequential derived lineages, the end points being denoted as the letters on the right side of the phylogeny. Numbers above the branches indicate the actual or estimated number of nucleotide substitutions that occurred along each branch, respectively. Actual substitutions were determined by sequencing 1091 base pairs of the ancestral viruses at each branch point in the tree. A striking example is the genomic study of microbes inhabiting our guts, our skin, the soil, and the oceans. Many of these microbes were not even known to science, since they could not be cultured in the lab or identified on the basis of morphology alone. First, regions of genes and genomes that do not change are likely of critical and unchanging function. This basic principle of molecular evolutionary conservation has pervaded all of biology, and has been a guiding principle underlying the study of function in the exponentially growing number of genome sequences now being completed for virtually every type of plant, animal, microbe, and virus. Conserved regions of genes are what have changed biology from being simply organism focused to drawing on comparative functional data about a gene of interest from all studied forms. Second, regions of molecules that do change have turned out to include both those of little or no function, and those for which rapid change is itself adaptive. Distinguishing between adaptive change and relaxed functional constraint can be challenging; however, numerous statistical and computational methods have been developed over the last two decades allowing discrimination between positively selected change and relaxed constraint, and an area of very active research is that of evaluating the functional and fitness consequences of the adaptive fixations. Another feature of molecular evolution that has emerged in the last decade is how much of the detected positive selection appears associated with conflict. Microbial and viral pathogens clearly have driven, and still drive, much of the rapid molecular evolution in genomes, but additional conflicts between males and females, between host genomes and transposable elements, and even between hosts and endosymbionts have emerged as important drivers of rapid molecular evolution, including the evolution of new genes and new gene functions. The role of noncoding portions of the genome in adaptation has also become strongly apparent in recent years. Variation in regulatory sequences (including enhancers, splicing machinery, and transcription factors) has been demonstrated as key to certain adaptive evolutionary changes in both molecules and phenotype (see chapters V. Clearly, much remains to be discovered about genomes and the means and mechanisms by which the molecules of life evolve and adapt. The changes in these molecules underlie the evolution of organismal form and function, and the field of molecular evolution is alive with new discoveries about how genomes evolve and how observed molecular changes contribute to the stunning biological diversity of life we observe around us. A solid introduction to many of the core principles of molecular evolution, though it was written in the "pre-genome" era and is thus missing many of the recent discoveries. The classic and well-written summary of the neutral theory argued by Motoo Kimura, a central figure in its development. A compre- hensive review of the development and use of the molecular clock concept from the 1960s through 2004. A recent summary of the patterns and mechanisms of the molecular evolution of genomes. A clear and concise summary of the statistical inference of natural selection from molecular data from within and between species. An introduction to molecular evolution and molecular population genetics with particularly clear figures and graphs. In eukaryotic organisms, a gene is expressed under the control of its own promoter and a combination of trans-acting factors that interact with other regulatory sequences. This is in contrast to prokaryotes, where a single promoter and set of regulatory sequences and few trans-acting factors dictate the coordinated expression of groups of linked genes. Processes that generate unequal outcomes for seemingly reciprocal mutational events. Cell size and nucleus size coevolve such that increased cell size corresponds with increased genome size. Genomic regions where crossovers occur at much higher rates than in other regions of the genome. Transposable elements multiply until they begin to affect (reduce) host fitness, thereby natural selection prevents their further proliferation. Throughout all forms of life, huge variation exists in the size and content of genomes, demonstrating the highly flexible, dynamic, and complex nature of their evolution. Processes including recombination and transposition of mobile genetic elements have been hypothesized as mechanisms for the expansion of eukaryotic nuclear genomes. Both adaptive and neutral processes have been implicated in the origin and evolution of these genomic elements, and understanding the nature of such mechanisms for genome evolution can provide important insights into the evolution of prokaryotic and eukaryotic diversity. Conditions that decrease the efficacy of selection make it more likely that novel elements such as introns, untranslated regions, and modularity in gene expression will become fixed in a population. Before the advent of high-throughput sequencing technologies and the resulting plethora of available genome sequence data, studies of genome evolution concentrated on comparing genome sizes across the tree of life. As such, prokaryotes and single-celled eukaryotes would possess fewer genes than multicellular eukaryotes; however, this relationship was not observed, and it was instead found that genome sizes ranged greatly, even within relatively closely related groups of taxa. Although a clear correlation between genome size and organismal complexity was not realized, two generalizable genome configurations are evident. Intergenic space in these organisms is limited, and blocks of genes-called operons-which largely encode for genes with functions in the same pathway or process, are cotranscribed using the same promoter and regulatory sequence(s). Second, in eukaryotes, genomes are dramatically larger and contained on one or more linear chromosomes. Thus, larger genomes contain proportionately fewer genes and more transposable elements than smaller genomes. If a specific sequence is required for increased recombination, that sequence should also be lost as a result of the very recombination that it induces. If a specific sequence is required for hot-spot activation, then a single base-pair change will inactivate it. Conversely, there are many sites in the genome that require a single base-pair change in order to become an activated hot spot; therefore, an evolutionary equilibrium may exist in which hot spots are degraded and introduced through these single base-pair changes. This explanation, coupled with the rapid evolution of hotspot activators like Prdm9, may explain the dynamic nature of genomic hot-spot locations even in very closely related organisms. Efficient repair of these breaks is critical, because their presence will disrupt replication and transcription. Errors in recombination can be devastating to an organism, but when they occur in the germ line, they also provide heritable restructuring events in genomes that contribute to genomic evolution in eukaryotes. Repetitive elements and self-replicating mobile elements are present throughout eukaryotic genomes. Because these elements have multiple homologous templates in the genome, recombination can potentially occur between any two, even if located on different chromosomes. When ectopic recombination occurs between these elements as a result of their sequence similarity, large-scale chromosomal rearrangements can occur, including sequence duplications, deletions, or inversions of large sections of chromosomes, and translocation of a chromosomal section from one chromosome to another. These changes can disrupt protein-coding sequences directly, as well as remove or add regulatory sequences that can result in aberrant expression of genes. Because of the inherent risk associated with the formation of these breaks, it is unsurprising that meiotic recombination appears a tightly regulated and evolutionarily constrained process. More unexpected are the constraints limiting the number of these breaks that result in a crossover event. Furthermore, these crossovers do not occur equally across the genome; rather, they are concentrated at hot spots, where rates of recombination are higher by several orders of magnitude than their flanking genome regions, or cold spots, in which crossover rates are extremely low. Organisms as closely related as humans and chimpanzees share no overlap in the genomic locations of hot spots, and intraspecific variation has even been observed within humans. Despite this fast rate of evolution of hot spots, there is mounting evidence that their location is sometimes dictated by specific sequence motifs. In the fission yeast Schizosaccharomyces pombe, several discrete sequences seven base pairs in length have been identified at active hot spots. In humans, one degenerate thirteen base-pair motif has been characterized at 41 percent of identified hot spots. Furthermore, in humans, the transcription factor Prdm9 is required for activation of these hot spots, and the amino acids that interact with the thirteen basepair motif are under strong positive selection. Generally, there are two classes of mobile elements characterized primarily by their mode of replication. Proliferation of these elements relies on the horizontal transfer of new elements from one organism to another, such as the transmission of small circular chromosomes containing the elements between prokaryotes. The second class of mobile elements is collectively referred to as retrotransposons. It is easy to see how the replication and insertion of mobile elements in a host genome could be slightly or strongly deleterious. For example, transposon insertion into a protein-coding region would most likely result in a frameshift, premature stop codon, or otherwise-aberrant protein sequence. Potentially, for this reason, a host has mechanisms to defend its genome from such elements. Because successful proliferation of a mobile element depends on the success of a host genome, it is also possible that transposable elements have built-in self-regulatory mechanisms preventing them from uncontrolled proliferation that would drive a host to extinction; however, such mechanisms have not been characterized. Importantly, as with all forms of mutation, mobile element insertion can on rare occasion give rise to evolutionary novelty. Because mobile elements encode their own machinery, multiple consequences can arise following their insertion into a new location. First, the elements contain protein-coding sequences and thus can introduce new coding regions into the genome (see chapter V. Second, these protein-coding sequences in mobile elements frequently have their own regulatory elements that can modify gene expression patterns of sequences, especially when adjacent to the insertion site. For example, the promoter region of a gene in the transposable element may recruit transcriptional machinery to a location near a host gene that has tight temporal or spatial regulation, causing it to be transcribed when it is normally silent. Indeed, it is hypothesized that centromeres and telomeres are often derived from mobile elements, and in some cases. Because of this increased mutation risk, it is not clear what, if any, advantage eukaryotes gain through the addition of 5? The mechanisms for evolution and origins of introns are much better understood than 5? In eukaryotes, introns in nuclear genes (spliceosomal introns) are processed by a nucleoprotein complex-the spliceosome- which is present in all eukaryotes and thus likely present in the most recent eukaryotic ancestor. This density of introns allows for a large number of potential transcripts per locus through alternative splicing, which in humans is responsible for the average 2. Although the current importance of introns is at least partly understood (alternative splicing, regulatory element content, etc. Debate over spliceosomal intron origin has been divided into two camps: those that propose the early evolution of introns prior to the divergence of eukaryotes and prokaryotes, and those that posit a later origin exclusively in eukaryotes. Whether spliceosomal introns arose early or late, there has been massive divergence in intron content in eukaryotes, making our understanding of the mechanisms underlying intron gain and loss of great importance. Because beneficial outcomes are extremely unlikely for the majority of transposable element insertions, adaptive hypotheses for the existence of these elements can be excluded for the most part. These elements are more frequently thought of as parasitic or selfish because of their lack of dependence on host machinery for replication, and their likely detrimental effects on host fitness. This hypothesis does not account for the role of other elements present in eukaryotic genomes, such as introns, and therefore cannot fully explain the increased genome size in eukaryotes. There are also several hypotheses for adaptive mechanisms underlying intron evolution in eukaryotes. Second, the presence of one or more introns allows for alternative splicing to occur, in which introns can be excised or retained, exons can be skipped, or exon length can vary depending on the usage of specific splice junctions. Further, the modular nature of genes that result from the inclusion of introns may have allowed for exon shuffling, in which mixing of domains from several different genes gives rise to genes with novel functions (see chapter V. Consistent with this, species that are intron poor have high conservation of their splice sites, whereas intronrich species have more degeneracy in their intron splice sites.
However milk allergy symptoms joint pain discount cyproheptadine 4mg, only one vestibular schwannoma case in 1253 patients was found to present with a basement syndrome allergy shots once a month cyproheptadine 4mg online. Thus allergy testing gold coast order 4mg cyproheptadine amex, the likelihood ratio for vestibular schwannoma given a basement syndrome was low (0 allergy medicine eye drops cyproheptadine 4 mg. Posterior Syndrome Posterior syndrome occurs when there is evidence for both abnormal horizontal canal or superior vestibular nerve and saccule or inferior vestibular nerve function allergy testing jersey uk cheap cyproheptadine 4 mg otc. To place this into context allergy medicine 10 month old discount cyproheptadine 4 mg free shipping, auditory evoked brain stem responses were estimated to have a likelihood ratio of 6 allergy symptoms summer cheap cyproheptadine 4mg visa. So being classified as having a global vestibular syndrome carried greater risk for vestibular schwannoma than an abnormal evoked potential study allergy medicine 906 purchase cyproheptadine 4 mg fast delivery. First, syndromes that reflect a superior vestibular nerve distribution are associated with complaints of vertigo. The syndrome approach mentioned here holds promise for improving our ability to explain and anticipate patient complaints. Ultimately, natural course of the problematic symptoms and fall risks will likely require both an understanding of the disease process underpinning the loss of function, and an understanding of what sensory functions are available, salvageable, and permanently lost. Key Points · Vestibular disorders can be based on the function of these three nerve branches- the superior vestibular, inferior vestibular, and cochlear eighth nerve branches. Global syndrome groups have a higher risk of vestibular schwannoma relative to other syndrome types. Medical treatments help with symptomatic care during acute vestibular crisis and have a role in prophylaxis. Surgery may help in those rare cases of perilymphatic fistula, superior canal dehiscence, or perhaps endolymphatic sac procedures. However, carefully developed, personalized treatments have been shown to substantially improve compensation. This relatively new area has begun to provide a mechanism for helping to treat the condition of patients who previously were told to live with their balance problem. Appropriate exercises prescribed by a physical therapist trained in vestibular rehabilitation can help speed and improve the recovery of many patients. Appreciating what structures are working, how they relate to subjective complaints and function deficits should ultimately help refine vestibular rehabilitation treatment plans even further. Careful history and physical examination, coupled with an understanding of vestibular anatomy, physiology, and pathophysiology, remain the bedrock for diagnosing problems of vertigo and imbalance. The electrophysiologic measures described in this chapter are merely extensions of the physical examination. Test data cannot be interpreted effectively without the same understanding of vestibular anatomy, physiology, and pathophysiology that underpins the initial history and physical examination. There are several key concepts for the clinician to keep in mind when measuring vestibular reflexes. First, there is a relationship between damaged sensory epithelia within the membranous labyrinth and abnormal reflexive behavior. When vestibulopathy is a possibility and the physical examination is unrevealing, the electrophysiological measures described in this chapter may be informative. The bilateral, bithermal caloric test provides a direct measure of vestibular weakness involving the horizontal canal and the superior vestibular nerve branch. Additionally, screening for positioning induced nystagmus is helpful in detecting benign paroxysmal positional vertigo. Visually guided eye movements may help characterize saccadic, pursuit, or optokinetic deficits that may also have been appreciated during the physical examination. Rotary chair testing is particularly useful in detecting bilateral vestibular weaknesses. Rotary chair test methods employ precisely controlled vestibular and visual stimuli, and are repeatable over time. In older patients, bilateral vestibular weakness may be missed on the standard caloric test. The ability to have a direct measure of utricular function is thus an important addition to the vestibular battery. Repeated measurements using an appropriate psychophysical method may be necessary for accurate measurements of ocular tilt using this method. These tests are not diagnostic in the sense that Vertigo and Balance 611 they point to specific sites of lesion. Rather, they measure behaviors that underpin normal control of upright stance in the functional sense. These tests reflect superior vestibular nerve, inferior vestibular nerve, and cochlear nerve branch function. Syndromes that involve the superior vestibular nerve branch tend to provoke stronger complaints of vertigo, and a tendency for benign paroxysmal positional vertigo. Syndromes that involve the inferior vestibular nerve branch are less likely to provoke strong complaints of vertigo and have a low risk for benign paroxysmal positional vertigo. Vestibular rehabilitation is an important addition to medical and surgical treatments for patients with complaints of vertigo and imbalance. Understanding how vestibular reflexes are impaired helps anticipate functional deficits. Evaluating the dizzy patient: Bedside examination and laboratory assessment of the vestibular system. The canalith repositioning procedure: For treatment of benign paroxysmal positional vertigo. Benign paroxysmal positional vertigo: 10-year experience in treating 592 patients with canalith repositioning procedure. Effectiveness of treatment techniques in 923 cases of benign paroxysmal positional vertigo. Cerebellar vermis lesions and tumours of the fourth ventricle in patients with positional and positioning vertigo and nystagmus. Positional and positioning nystagmus in healthy subjects under videonystagmoscopy. The relationship between falls history and computerized dynamic posturography in persons with balance and vestibular disorders. A new set of criteria for evaluating malingering in workrelated vestibular injury. Dehiscence of bone overlying the superior canal as a cause of apparent conductive hearing loss. Characteristics and clinical applications of vestibularevoked myogenic potentials. Comparison of the head elevation versus rotation methods in eliciting vestibular evoked myogenic potentials. Autonomic dysfunction has important implications for health and disease yet is clinically under recognized. Clinical signs of autonomic dysfunction are easily overlooked, and neural activity in the autonomic nervous system is difficult to record directly. Although sympathetic nerve function in peripheral nerves can be recorded with fine-tipped tungsten electrodes, this technique is difficult to apply clinically. Therefore, the assessment of autonomic function depends primarily on measuring the response of the autonomic nervous system to external stimuli. The measurements of sweating (Chapters 36 and 38), cardiovascular activity and peripheral blood flow (Chapters 37 and 39), and central autonomic-mediated reflexes provide insight into the broad range of disorders that affect the central and peripheral components of the autonomic nervous system-from the hypothalamus to the autonomic axons in the trunk and limbs. With better understanding of the clinical importance of measuring autonomic function and with increasing use of newly available tests of cardiovagal function, segmental sympathetic reflexes, postural hemodynamics, and power spectral analysis, the tests and measurements of autonomic function will be of greater benefit in patient care. Pain is mediated mainly through small nerve fibers, particularly in the autonomic nervous system. Measurements of their function can help elucidate the mechanisms underlying pain, especially peripheral pain. The emerging modalities for assessment of pain pathways include quantitative sensory tests, autonomic tests, microneurography, and laserevoked potentials (Chapter 40). Direct recording of spontaneous electric activity in nerves by microneurography is tedious but can be particularly helpful. Its integrative functions coordinate input from peripheral and visceral afferent nerves to orchestrate a dynamic balance among organ systems. Its adaptive functions react moment by moment to the various forms of stress the body experiences. It consists of sympathetic (thoracolumbar) and parasympathetic (craniosacral) divisions. The enteric nervous system, located in the wall of the gut, is considered a third division of the autonomic nervous system. The autonomic nervous system thus regulates and coordinates such physiological functions as blood pressure and heart rate, respiration, body temperature, sweating, lacrimation, nasal secretion, pupillary size, gastrointestinal motility, urinary bladder contraction, sexual physiology, and blood flow to the skin and many organs. Autonomic neuropathies that disconnect central autonomic centers and autonomic ganglia from their peripheral effectors 617 618 Clinical Neurophysiology may result in deficits in autonomic function. Examples include orthostatic hypotension due to adrenergic failure, heat intolerance due to sudomotor failure, gastroparesis, hypotonic bladder, and erectile failure. Autonomic centers disconnected from inhibitory influences may give rise to episodic autonomic hyperfunction. Examples include autonomic dysreflexia and hypertonic bladder following spinal cord trauma, diencephalic syndrome following head injury, hypertensive surges of baroreflex failure following irradiation to the carotid sinuses, auriculotemporal syndrome, and catecholamine storms in pheochromocytoma. Autonomic disturbances frequently accompany neurologic illnesses affecting motor or sensory systems or may occur in isolation. More frequently, accurate characterization, localization, and grading of autonomic dysfunction require a careful history to elicit subtle symptoms, a neurological examination attentive to autonomic signs, and testing in a clinical autonomic laboratory. The differential diagnosis of autonomic failure includes the peripheral neuropathies (many of which involve autonomic fibers), central degenerative disorders, and medical disorders that impact the autonomic nervous system. Peripheral autonomic failure frequently occurs in small fiber neuropathies, particularly in diabetes mellitus and amyloidosis. Autonomic disturbances are common also in botulism, diphtheritic neuropathy, and Chagas disease. The presence of autonomic failure has important implications for clinical management as well as for prognosis. Disruption of autonomic function can influence the longterm risks of morbidity,1, 2 mortality,35 and intraoperative mortality. Unlike the easily reproducible function of somatic motor or sensory nerves, autonomic nerve function is difficult to evaluate precisely in humans. In general, evaluation of autonomic function has been restricted to noninvasive recordings of heart rate, blood pressure, blood flow, or sweat production. The interpretation of the results of these tests may be difficult, because (1) the effector organs react slowly to variations in neural input, (2) the interactions of sympathetic and parasympathetic outputs at a single target level are complex, and (3) autonomic responses are affected by pharmacologic, hormonal, local chemical, and mechanical influences. This chapter provides an overview of some aspects of autonomic function that may help with interpreting the results of noninvasive autonomic tests commonly used clinically. Purpose and Role of Autonomic Testing · Recognize the presence, distribution, and severity of autonomic dysfunction. Quantitatively evaluate autonomic dysfunction over time To define the progression or remission of autonomic disease To assess the response to therapy. Visceral Afferents Visceral receptors generally are slowly adapting mechanoreceptors or chemoreceptors that have a low level of spontaneous activity and are innervated by small myelinated and unmyelinated fibers. Organization of the sympathetic and parasympathetic outputs of the autonomic nervous system. This reflects important differences in the functional organization of autonomic and somatic efferents. Autonomic output involves a two-neuron pathway that has at least one synapse in an autonomic ganglion. At most sympathetic neuroeffector junctions the primary postganglionic neurotransmitter is norepinephrine, which acts on the various subtypes of - and -adrenergic receptors. The primary postganglionic neurotransmitter at all parasympathetic neuroeffector junctions is acetylcholine, which acts on the various subtypes of muscarinic receptors. Whereas the main consequences of denervation in the striated muscle are paralysis and atrophy, postganglionic efferent denervation produces an exaggerated response of the target when it is exposed to the neurotransmitter. This phenomenon, called denervation supersensitivity, is evidence of a lesion involving postganglionic neurons in which the remaining neuroeffector receptors are upregulated. Activity of most autonomic effectors is modulated by dual, continuous sympathetic, and parasympathetic influences. It predominates in control of the salivary glands, sinoatrial node, gastrointestinal tract, and bladder. The sympathetic and parasympathetic systems may interact at several levels, including that of the central nervous system, autonomic ganglia, neuroeffector junction, and target organ. Postganglionic autonomic neurons send unmyelinated (type C) axons to innervate peripheral organs. At most sympathetic neuroeffector junctions the primary postganglionic neurotransmitter is norepinephrine. In the sweat glands, sympathetic effects are mediated principally by acetylcholine. The primary postganglionic neurotransmitter at all parasympathetic neuroeffector junctions is acetylcholine. Sympathetic functional units include skin vasomotor, muscle vasomotor, visceromotor, pilomotor, and sudomotor units. Their postganglionic fibers destined for the trunk and limbs follow the course of spinal nerves or blood vessels or both. Spinal fibers join the peripheral spinal (somatic) nerve through the gray ramus communicans. These fibers provide vasomotor, sudomotor, and pilomotor input to the extremities and trunk. Sympathetic fibers are intermingled with somatic motor and sensory fibers, and their distribution is similar to that of the corresponding somatic nerve. Most sympathetic fibers are destined for the hand and foot and are carried mainly by the median, peroneal, and tibial nerves and, to a lesser extent, the ulnar nerve. For example, nociceptive stimuli reflexively activate segmental circuitry that generates excitation of vasoconstrictor outflow to skeletal muscle and inhibition of vasoconstrictor outflow to the skin. For example, during execution of the cold pressor test, decreases in cutaneous blood flow in an arm exposed to ice cold water are accompanied by cutaneous vasoconstriction in the contralateral forearm. All segmental spinal reflexes are subject to supraspinal modulation through several parallel pathways arising in the hypothalamus, pons, and medulla and innervating the sympathetic preganglionic neurons. Indirect methods include noninvasive tests of sudomotor and cardiovascular function described in Chapters 3639; measurement of plasma norepinephrine concentration in forearm veins with the subject supine and standing;18, 19 assessment of splanchnic20, 21 and cerebral blood flow22, 23 using Doppler techniques; and assessment of sympathetic innervation of the heart using radioisotope methods. This technique allows multiunit recordings of two different types of outflow: skin sympathetic nerve activity and muscle sympathetic nerve activity. Postganglionic sympathetic fibers travel with spinal motor and sensory fibers or with arteries to provide vasomotor, sudomotor, and pilomotor innervation to the trunk and limbs. Most sympathetic fibers supplying the foot are carried by the peroneal and tibial nerves. Unlike somatic motor neurons, sympathetic preganglionic neurons are not monosynaptically driven by afferent input. Sympathetic reflexes are segmentally biased, predominantly uncrossed, and exhibit ipsilateral, function-specific, reciprocal and nonreciprocal patterns of response. Sympathetic function can be assessed noninvasively by indirect techniques, or invasively by direct microneurographic recording of postganglionic sympathetic nerve activity. Decreased or increased environmental temperature can produce selective activation of the vasoconstrictor or sudomotor system, respectively, with suppression of activity in the other system. Emotional stimuli or inspiratory gasp also increases spontaneous skin sympathetic activity, but in this case the bursts are caused by simultaneous activation of sudomotor and vasomotor impulses. Arteriovenous skin blood flow is carried by low-resistance arteriovenous shunts, which receive abundant sympathetic vasoconstrictor input and have a key role in thermoregulation. Skin blood flow is also controlled by somatosympathetic reflexes13, 14 and three local axon reflexes: (1) the axon flare response, (2) the sudomotor axon reflex, and (3) the venoarteriolar reflex. The venoarteriolar reflex is mediated by sympathetic vasomotor axons innervating small veins and arterioles. Skin vasomotor activity has been studied clinically by using several noninvasive methods for measuring skin blood flow, including plethysmography and laser Doppler flowmetry. The segmental pattern of distribution of sudomotor fibers to the trunk and limbs is irregular and varies substantially among individuals and even between the right and the left sides of an individual. Key Points · Sympathetic vasomotor, pilomotor, and sudomotor innervation of skin effectors has primarily a thermoregulatory function. However, this activity in the arm and leg can be dissociated during mental stress and during forearm ischemia after isometric exercise. The respiratory cycle, changes of posture, or the Valsalva maneuver may modulate the muscle sympathetic activity caused by changes in arterial pressure. However, hypercapnia, hypoxia, isometric handgrip, emotional stress, or the cold pressor test increase muscle sympathetic activity despite unchanged or increased arterial pressure. Parasympathetic control is provided by cardiovagal neurons in the nucleus ambiguus and dorsal motor nucleus in the medulla. Effects mediated by the vagus nerve have a shorter latency and duration than those mediated by the sympathetic nerves. Heart rate has spontaneous fluctuations, which reflect changing levels of autonomic activity modulating sinus-node discharge. Heart rate variability is correlated inversely with age in normal subjects at rest. Upright posture in humans dramatically increases sympathetic nerve activity and produces a large increase in low-frequency heart rate power. The primary role of arterial baroreflexes is the rapid adjustment of arterial pressure around the existing mean arterial pressure.
In contrast allergy symptoms red ears discount 4mg cyproheptadine free shipping, a tree with a high likelihood is a better estimate allergy shots for dust mites purchase cyproheptadine 4 mg fast delivery, and the tree that results in the maximum likelihood value among all trees is viewed as the best estimate of the tree allergy treatment 3 phases effective 4mg cyproheptadine. The probability statements for a data pattern can be constructed by considering the probability of all possible character state changes across a single branch in the tree (the transition probabilities for the branch) allergy testing gluten cheap 4 mg cyproheptadine overnight delivery. By assuming that evolutionary events on different branches are independent allergy treatment sydney buy 4mg cyproheptadine with visa, one can combine per-branch transition probabilities into probabilities for the evolutionary history of a character across the entire tree allergy symptoms before period generic 4mg cyproheptadine. It is often convenient to consider the rates at which different changes could occur allergy medicine for 6 month old baby purchase 4mg cyproheptadine. Mathematical transformations allow us to extrapolate the effects of an evolutionary process occurring at a certain rate over any timescale of interest allergy medicine 773 purchase 4mg cyproheptadine amex. After a description of character evolution is formulated in terms of rates of change, it is possible to calculate a likelihood for any combination of tree topology and branch lengths. By maximizing the likelihood we can find the parameter values that match the data most closely. One implication of treating branch lengths as unknown parameters is that every character we observe in a data matrix provides information. An entirely constant character will not "directly" prefer one tree over another, but it will provide evidence that branch lengths are short. This could have an indirect effect on which tree best fits the data, so even constant characters can alter tree inference. Relying on a summary of the data (the distance matrix) rather than the full data should make distance methods less powerful than character-based methods. In general, no compelling statistical reasons have been advanced for preferring distance-based methods over likelihood-based approaches. Distance-based approaches continue to be widely used, however, because they provide reasonable estimates of the tree very quickly even for very large data sets. Our models of character evolution are dramatic oversimplifications of the real evolutionary processes. Distance-based methods for tree reconstruction simplify the tree inference problem by trying to explain only the observed divergences between the tips of the tree. A tree makes a prediction about the evolutionary distance (the number of character state changes that have occurred) between each pair of tips of the tree. We can observe pairwise divergence in the characters that we study, so we have an empirical estimate of the tip-to-tip distance. From the character matrix, we can calculate a divergence between each taxon to every other taxon and summarize these calculations in a taxon-by-taxon distance matrix. The combination of tree topology and branch lengths with tip-to-tip divergences closest to the observed distance matrix is judged to be the best estimate of phylogeny. Distance-based approaches treat the distance matrix as if it were the only data relevant to tree inference. Because distance methods do not have to "map" evolutionary events on the tree for each character, they can be very fast. Condensing a character matrix into a distance matrix implies a loss of information. When we compare characters between two taxa (tips on a tree) the number of differences represents a minimum number of evolutionary events that must have occurred. Thus, the observed pairwise distance matrix is not an error-free representation of the evolutionary distance between tips. Models can be used to correct the pairwise distance estimates for repeated changes at the same position (the "multiple hits" problem). But even a corrected pairwise distance is often an imprecise estimate of the preceding sections have focused on the statistical basis of estimating a tree, specifically on the correspondence between various estimation methods and different ways of assessing the fit between a phylogenetic hypothesis and the observed data. Developing the computational machinery to conduct phylogenetic inference is a complementary, and very active, area of research. The number of possible trees is enormous, so scoring every possible tree is not feasible. In general, software for phylogenetic estimation works by generating a rough initial solution, then trying to improve the estimate by looking at similar trees. A procedure called stepwise addition builds up a tree estimate by adding taxa to a growing tree one at a time. Placements made in early steps may be suboptimal when new data are added to the tree. If the perturbation results in a tree with a better score, we have improved our solution, and we can continue searching for a better tree. If we try a large number of perturbations and fail to find a tree with a better score, we can terminate the search. The final tree will be a good approximation of the tree with the optimal score even if we cannot guarantee that our search found the best tree. Repeatedly performing searches from different starting points can reveal whether this type of hill-climbing approach appears to be working on a particular data set. If each starting point yields a different final tree, then the landscape of tree scores is very complex and there is a good chance that none of the searches identified the "global" optimum. Many variations of this general strategy of tree searching have been studied, and it is now feasible to reconstruct trees of hundreds and even thousands of taxa. When dealing with large data sets, one can rarely Phylogenetic Inference be confident that the optimal tree has been found; however, it is unlikely that one could reconstruct a huge tree with no error at all. The crucial question becomes, "What aspects of the tree are strongly supported by the data? Strongly supported branches in a tree are usually easy to find during tree searching, so most phylogenetic analyses are limited more by the amount of information in the data rather than by the efficiency of tree-searching software. Explicit criteria can help us choose among a set of alternative families of models, and formal tests of model adequacy can identify cases in which our inferential models are clearly unrealistic. Even with a satisfactory model of the evolutionary processes generating the data, we still must acknowledge the possibility that limitations in our data can lead to an incorrect estimate. Given a clade in our estimate of phylogeny, such as the grouping of human with orangutan, we would like to know whether the grouping could simply be the result of sampling error rather than a true evolutionary signal. Sampling error refers to the mistakes in estimation caused by a small sample of data. A common approach in statistics is to calculate a P value to evaluate the strength of evidence about a proposition. Roughly speaking, a P value is the probability of seeing at least as much evidence against a proposition even if the proposition is true. It helps us assess whether it is plausible to discount our result as merely an artifact of sampling error. To calculate a P value for a clade within a phylogeny, we quantify the support for the group in a numerical statistic. The most appealing choice is the difference in score between the best-scoring tree (our estimate) and the best tree that does not contain the clade of interest. For example, in the primate mitochondrial genome example, the tree with the best parsimony score grouped human and orangutan; this tree required 7990 changes to explain the data. The best alternative tree places orangutan with the baboon; that tree required 8324 steps to explain the sequence data; thus, we can say that the human + orangutan tree is 334 changes better than the next-best tree. Obtaining a large difference in scores clearly implies that we have more compelling evidence. We might want to calculate a P value for the hypothesis that human and orangutan are not close relatives. To do this we must answer the question, "If human and orangutan were not a true group on this phylogeny and we randomly sampled 16,767 sites, what is the probability that we would obtain a data set that yields a score difference of at least 334 steps in favor of an incorrect tree? Phylogenetic trees are difficult parameters to deal with, and we do not have the convenience of calculating a simple number for a tree and looking it up in a standard statistical table; nevertheless, we can still apply the core insights of statistical testing and identify groupings in a phylogenetic estimate that are weakly supported and likely to be overturned by subsequent analyses. If we think our data are very "clean" (unlikely to generate patterns that support spurious groupings), then it seems very unlikely that we would see a score difference of 334 entirely from sampling error. If our data appear to have lots of homoplasy, we might see this much-erroneous signal. We can look at the number of sites that supported different groupings in the original data set to see how variable the score would be as a result of resampling. Alternatively, we could use a computer to simulate the effect of sampling error by generating many artificial data sets. By counting the proportion of simulated data sets that display at least 334 steps of support for spurious groupings, we can approximate a P value. Bootstrapping is the most common way to assess the effect of sampling error on tree inference. In bootstrapping, we create many "pseudoreplicates" of our original data by randomly sampling from the pool of characters we observed. In each pseudoreplicate, a different set of characters will be overrepresented and another set will be excluded. By conducting phylogenetic analysis on each of these pseudoreplicates, we can discover which groupings in the tree are sensitive to sampling error. The bootstrap proportion for a group is the proportion of pseudoreplicate analyses that supported the group. Bootstrapping is computationally demanding, because hundreds of tree searches must be performed, and it does not directly yield a P value; however, it does provide a useful summary of clades that are well supported. Bayesian approaches formalize the ways we can use data to update our beliefs about the world. We start by considering all parameter values, assigning each parameter value a prior probability. For instance, in the case of the primate tree, a person who had never studied anthropology or mammalogy might be completely uncertain about whether the correct 66 Phylogenetics and the History of Life then we can express that as a prior probability statement and use that information in a Bayesian framework. In the vast majority of cases, Bayesian phylogenetic inference is conducted by a computer-simulated walk through the space of all parameter values. We can design the rules for the simulation in a very specific way so the walk tends to avoid parameter values with low likelihood (or lower prior probability). Running these simulations for a large number of iterations provides a set of parameters that are sampled in proportion to their posterior probability. This Markov chain Monte Carlo simulation approach is an elegant solution to the difficulties of exploring a large parameter space, but it requires considerable care to ensure it provides reliable results. The Phylogenetic Handbook: A Practical Approach to Phylogenetic Analysis and Hypothesis Testing. An advanced book covering theory and descriptions of software for tree estimation. Maximum likelihood as an alternative to parsimony for inferring phylogeny using nucleotide sequence data. In such a case, the person might assign each tree a prior probability of 1/3 to reflect the fact that he thinks each scenario is equally likely (and the sum of probabilities must be 1). Technically speaking, this prior probability for the tree topology is actually an integral of probability densities over all the possible branch-length combinations for the tree. Before looking at the data, few people would be confident about specifying a reasonable branch length, but some combinations might seem implausible. For example, we might be surprised if the branch leading to human were thousands of times longer than the branch leading to its closest relative; by assigning such combinations low prior probability, one is able to bring previously learned insights to bear on an analysis. This step is not subjective at all; in fact, it is simply an exercise in applying the rules of probability. One very attractive feature of Bayesian inference is its ability to produce a single-best estimate of a parameter (for instance, the phylogeny with the highest posterior probability), but also an easily interpreted statement of support. If the posterior probability for a tree or a clade is close to 1, then those aspects of evolutionary history are strongly supported. The result of a Bayesian analysis is a summary that blends any prior knowledge with information from the data. In practice, we often have only vague knowledge of the model before seeing the data, so the effect of the likelihood dominates the inference. The molecular evolutionary clock Molecular clock dating Testing the molecular clock Statistical methods for divergence time estimation Maximum likelihood estimation of divergence times Bayesian estimation of divergence times Fossil calibrations Relaxed clocks and prior model of rate drift Perspectives this chapter reviews the history of the molecular clock, its impact on molecular evolution, and the controversies surrounding mechanisms of evolutionary rate variation and the application of the clock to date species divergences. We review current molecular clock dating methods, including maximum likelihood and Bayesian methods, with an emphasis on relaxing the clock and on incorporating uncertainties into fossil calibrations. A "relaxed" clock model allows rates to vary across lineages in an orderly way; there may be a "local clock" with constant rates in subsets of species (in a likelihood analysis), or there may be lineage-specific rates that are either independent observations from a common distribution or correlated between ancestral and descendant species (in a Bayesian analysis). One of the first methods for modeling sequence substitution rate evolution among lineages (a relaxed molecular clock). This early heuristic procedure penalizes changes in rate between ancestral and descendant branches while maximizing the probability of the data. In the simplest case, an interior node on the tree is assigned a fixed age, and a molecular clock is then applied in an analysis of the sequence data to estimate the absolute ages of the remaining nodes. More sophisticated calibration methods use Bayesian methodology to accommodate uncertainties in the fossil record, by specifying a distribution for a node age (instead of a fixed constant). A regression-based method for determining how much remaining uncertainty for node ages is due to uncertainties in fossil calibration times (or lack thereof) and how much to insufficient sequence data. In the early 1960s, it was observed that the amino acid differences between aligned hemoglobin or cytochrome c sequences from different species were roughly proportional to the times of divergence between the species (according to the fossil record). These observations led Emile Zuckerkandl and Linus Pauling to propose the hypothesis of a molecular evolutionary clock in 1965. The clock was envisaged as a stochastic one, with "ticks" corresponding to nucleotide or amino acid substitutions, which occur at random time intervals. Although particular substitutions occur at random times, the rate at which substitutions occur is assumed to be constant or "clocklike" through time and across lineages. The process is analogous to the way in which the random decay of isotopes can be used to construct an atomic clock. Furthermore, much the way that different isotopes have a characteristic rate of radioactive decay, different proteins can have different evolutionary rates, meaning that their molecular clocks tick at different rates. Perhaps because of the generation time effect or other correlated life history variables, for example metabolic rate, substitution rates tend to be negatively related to body size, with high rates in rodents, intermediate rates in primates, and slow rates in whales. Species with small body sizes tend to have shorter generation times and higher metabolic rates. The negative correlation between substitution rate and body size has been supported in some studies but questioned in others. The reliability of the clock and its implications for the mechanism of molecular evolution were a focus of immediate controversy. The molecular clock hypothesis was proposed at a time when the neo-Darwinian theory of evolution was generally accepted by evolutionary biologists, according to which the evolutionary process is dominated by natural selection. A constant rate of evolution among species as different as mice and monkeys was incompatible with that theory. Species living in different habitats, with different life histories and generation times, must be under very different regimes of selection (and therefore should have different substitution rates). When the neutral theory of molecular evolution was first proposed (by Motoo Kimura in 1968 and by Jack King and Thomas Jukes in 1969), the observed clocklike behavior of molecular evolution was considered major supporting evidence. The neutral theory emphasizes random fixation of neutral or nearly neutral mutations (see chapter V. Under such a model, the rate of substitution is equal to the neutral mutation rate, independent of factors such as environmental change and population size variation. If the mutation rate is similar and the function of a protein remains the same across species (so that the same proportion of mutations are neutral), a constant substitution rate is expected. Rate differences among proteins are explained by the presupposition that different proteins are under different functional constraints, with a different proportion of amino acids experiencing neutral mutations. The neutral theory is not the only mechanism compatible with clocklike evolution; neither does the neutral theory always predict a molecular clock. Under the clock assumption, the expected distance between sequences increases linearly with time of divergence. When external information about the geological ages of one or more divergence events on a phylogeny is available, based on the fossil record or certain geological events, the distances between sequences or the branch lengths on the tree can be converted into absolute geological times. The earliest application of the clock to estimate divergence times was by Zuckerkandl and Pauling in 1962, who used an approximate clock to date duplication events among a, b, g, and d globins of the hemoglobin family. The outcomes of molecular clock analyses have often produced controversies, usually because the molecular dates are at odds with the fossil record. Fossil forms of metazoan phyla appear as an "explosion" around 540 million years ago in the early Cambrian, but most molecular estimates of the ages of these divergence events have been much older, sometimes twice as old. Part of the discrepancy between molecular and fossil data is due to the incompleteness of the fossil record. Fossils provide information concerning the date by which Molecular Clock Dating a newly diverging lineage had developed diagnostic morphological characters. There may be a lag between the time that a lineage arose and the age of the first fossil with the derived traits of the descendants. Molecular dating, in contrast, infers ages of nodes (divergence events among ancestral lineages) in a phylogenetic tree. Fossil-based dates therefore tend to be younger than those derived from molecular data. Another source of discrepancy can be inaccuracies and deficiencies in molecular time estimation. Despite sometimes acrimonious controversies, the interactions between molecules and fossils have been a driving force in this research area, since they have prompted reinterpretations of fossils, critical evaluations of molecular dating techniques, and the development of more advanced analytical methods. Our focus in this chapter is on statistical methods for testing the clock hypothesis, and on likelihood and Bayesian methods for dating species divergence events under global and local clock models. In such analyses, fossils are used to calibrate the clock, that is, to translate sequence distances into absolute geological times and substitution rates. One can use the dates at which particular viruses were isolated to calibrate the clock and to estimate divergence times, using essentially the same techniques as discussed here. Indeed, such dated viral sequences are sometimes referred to as "fossil sequences," although most such samples were isolated during the last 100 years and are not true fossils.
With increasing time since the split allergy symptoms gas purchase cyproheptadine 4 mg visa, genetic divergence accumulates allergy shots nhs cheap cyproheptadine 4mg, and diagnosing the separate lineages becomes easier allergy shots to cats purchase 4mg cyproheptadine with mastercard. Unlike the fst definition of race allergy symptoms brain fog generic 4 mg cyproheptadine fast delivery, no arbitrary threshold of differentiation is set a priori allergy testing asthma discount cyproheptadine 4 mg on-line. A split into separate lineages also means that the genetic differences among the races would define an evolutionary tree analogous to an evolutionary tree of species allergy medicine you can take while pregnant 4mg cyproheptadine amex. Statistical methods exist for testing the null hypothesis that the genetic variation within a species has a treelike structure sulfite allergy symptoms uk purchase cyproheptadine 4 mg on-line, and other statistics test the null hypothesis that the entire sample defines a single evolutionary lineage allergy symptoms weather changes 4 mg cyproheptadine for sale. Therefore, just as with the fst definition, the lineage definition of race can be implemented for all species in an objective fashion using uniform criteria, thereby avoiding a human-specific or cultural definition of race. Obviously, the culturally defined racial categories of "white" and "black" do not have the same genetic meanings in the United States and Brazil. It is clear that an objective, culture-free definition of race is required before the question about the existence of biological races can be answered. One way of ensuring a culture-free definition of race is to use a definition that is applied to species other than humans. The word race is not commonly used in the nonhuman literature; instead, the word subspecies is used to indicate the major types or subdivisions within a species. We will apply these culture-free definitions to humans to avoid an anthropocentric definition of race. Biologically, races are geographically circumscribed populations within a species that have sharp boundaries that separate them from the remainder of the species. In traditional taxonomic studies, the boundaries were defined by morphological differences, but increasingly these boundaries are defined in terms of genetic differences that can be scored in an objective fashion in all species. Most local populations within a species show some degree of genetic differentiation from other local populations by having either some unique alleles or different frequencies of alleles. If every genetically distinguishable population were elevated to the status of race, then most species would have hundreds to tens of thousands of races. This would make the concept of race nothing more than a synonym for a local population. There is a consensus that race or subspecies should refer to a degree or type of genetic differentiation that is well above the level of genetic differences that exist among local populations. Both quantitative and qualitative criteria are used to define these racial genetic boundaries. Quantitatively, one commonly used threshold is that two populations with sharp boundaries are considered to be different races if 25 percent or more of the genetic variability that they collectively share is found as between- Evolution and Notions of Human Race 2. Genetic differentiation among populations of Before addressing the existence of human races, we first apply these definitions of race to our closest evolutionary relative, the chimpanzee. In this manner, the definitions can be applied in a context that avoids the emotion and cultural biases that inevitably creep into discussions of human race. Based on morphological differences, the common chimpanzee (Pan troglodytes) has been subdivided into five races or subspecies: P. Gonder and coworkers (2011) genetically surveyed chimpanzees throughout their range. They discovered sharp genetic differences separating the Upper Guinea and Gulf of Guinea populations from all other populations, but with less sharp genetic boundaries between the equatorial African populations. The Upper Guinea and Gulf of Guinea populations are above the 25 percent threshold for contrasts with each other and with all other chimpanzee populations. However, the three regions sampled in equatorial Africa are all well below the 25 percent threshold. Hence, there are three races or subspecies of common chimpanzees using the threshold criterion: P. If chimpanzees are subdivided into separate evolutionary lineages, the genetic differences among lineages should define a treelike structure characterized by splits and isolation. There are genetic differences between different geographic areas (table 1), but such genetic differentiation can also arise when gene flow (genetic interchange associated with individuals who disperse from their birth population) occurs but is restricted by geography. For example, gene flow can be restricted when most dispersal is limited to nearby local populations. Such stepping-stone models yield a pattern of isolation by distance in which the degree of genetic differentiation between two populations increases with increasing geographic distance between them. Genetic differentiation structured by isolation by distance can be distinguished from genetic differentiation due to lineage splits by testing for constraints on genetic distances. Note: Rst is related to fst but incorporates a mutational model for microsatellites. Under isolation by distance, the genetic distance (measured, say, by the fst value between a pair of populations) should increase with increasing geographic distance; that is, the fst between A and B should be less than the fst between A and C. In contrast, suppose populations A, B, and C represent separate evolutionary lineages (races) such that A split from the common ancestral population of B and C in the past, followed by a more recent split between populations B and C. This results in an evolutionary tree of populations such that genetic distance between populations increases with the time since their split from a common ancestral population. In this hypothetical case, the genetic distances between populations A and B and between populations A and C should be the same, since they both involve a split from the same ancestral population. Hence, the expected pattern of genetic distances differs for trees versus isolation by distance, and formal statistical tests exist to determine whether the pattern of genetic differentiation is consistent with the special constraints imposed by an evolutionary tree. Another method for testing for a treelike structure is based on finer geographic sampling. As more sites are sampled under an isolation-by-distance model, the geographically intermediate populations should also have intermediate genetic distances. In contrast, when the populations are grouped into a smaller number of evolutionary lineages, genetic distances among populations within a lineage should be relatively small, although they may show an isolation-by-distance pattern within the geographic range occupied by a particular lineage. However, the genetic distances are expected to show a large, sudden increase when the geographic boundary between two lineages is crossed. When the chimpanzee genetic data are used to estimate an evolutionary tree of populations, the resulting 804 Evolution and Modern Society Table 2. This tree predicts that the Upper Guinea population should be equally distant from all the other populations, and table 1 shows that this prediction is supported when the error in estimating the distances is taken into account. This tree also predicts that the Gulf of Guinea population should be equally distant from all the equatorial African populations but that this distance should be smaller (less time since the split) than the distances involving the Upper Guinea population. However, the genetic distances among the three equatorial African populations show the isolation-by-distance pattern on an east-west axis. Hence, chimpanzees do show a treelike structure of genetic differentiation with three lineages: Upper Guinea, Gulf of Guinea, and the combined equatorial African populations. Hence, races do exist in chimpanzees under the lineage definition, and they correspond exactly to the same three races defined by the quantitative threshold definition of race. They used a computer program to sort individuals or portions of their genomes into five groups and discovered that the genetic ancestry of most individuals was inferred to come mostly from just one group. Moreover, the groups corresponded to five major geographic populations: (1) sub-Saharan Africans; (2) Europeans, Near and Middle Easterners, and Central Asians; (3) East Asians; (4) Pacific populations; and (5) Amerindians. This paper was the most widely cited article from the journal Science in 2002, and many of these citations claimed that this papersupportedtheideathatraceswerebiologicallyrealin humans. When they increased the number of groups beyond five, they also obtained an excellent classification into smaller, more regional groups. Hence, they showed that with enough genetic markers, it is possible to discriminate most local populations from one another. Recall that genetic differentiation is necessary but not sufficient to define races, so even if there is a consensus that five groups is the right number, genetic discrimination alone does not necessarily mean that these five groups are races. Assuming for now that the five groups are the meaningful populations, do these groups satisfy the quantitative threshold definition of race? Table 2 shows how the genetic variation is hierarchically partitioned into differences among individuals within the same local population, differences among local populations within the same "race," and among "races. In contrast with chimpanzee races, the five major "races" of humans account for only 4. The genetic variation in our species is overwhelmingly variation among individuals (93. As for the lineage definition, a treelike structure of genetic differentiation has been strongly rejected for every human data set subjected to testing for the constraints expected from an evolutionary tree of populations. When Rosenberg and coworkers published their results in 2002, their geographic sampling was coarse. It is now known that the computer program used in these studies generates well-differentiated populations as an artifact of coarse sampling from species characterized by isolation by distance. Figure 1 shows a plot of the pairwise fst values of Evolution and Notions of Human Race 805 0. Consequently, it is not surprising that when Behar and coworkers (2010) sampled Old World populations more finely and used the same computer program used in the 2002 study, most individuals showed significant genetic inputs from two or more populations, indicating that most human individuals have mixed ancestries. The "races" so apparent to many who cited Rosenberg and coworkers simply disappeared with better sampling. These results and figure 1 falsify the hypothesis that humans are subdivided into evolutionary lineages. The distinct genetic states that exist in such regions (called haplotypes) reflect the accumulation of various mutations during evolutionary history. This evolutionary history, called a haplotype tree, is the history of the genetic variation in that genomic region and is not necessarily the history of the populations that bear this variation. Indeed, if a species has sufficient gene flow, there can be no evolutionary tree of populations because there are no population splits; however, there will still be haplotype trees for each nonrecombining region of the genome. Haplotype distributions can be influenced by population-level history, but the population-level information embedded in a haplotype tree must be extracted carefully. It is never justified to equate a haplotype tree directly to an evolutionary history of populations. The oldest inferred event is an out-ofAfrica range expansion into Eurasia genetically dated to about 1. The paleoclimatic data indicate that the Sahara region experienced repeated minor wet periods such that the Sahara is unlikely to have been a dispersal barrier on a timescale of tens of thousands of years. The null hypothesis of complete genetic isolation during the Pleistocene is decisively rejected. Consequently, even during the Pleistocene, Old World human populations were not subdivided into isolated and independently evolving lineages. The next major event shown in figure 2 is a second population expansion out of Africa into Eurasia around 700,000 years ago, corresponding to the spread of the Acheulean tool culture out of Africa into Eurasia during the second major Saharan wet period of the Pleistocene. The null hypothesis of no admixture between the expanding population and the Eurasian populations is rejected. Hence, the Acheulean expansion was marked by 806 Evolution and Modern Society Time Africa South Europe North Europe South Asia North Asia Pacific Americas Gene flow with isolation by distance and some long-distance dispersal Range expansions into new areas Male-mediated out-of-Asia expansion Out-of-Africa expansion of Homo sapiens 0. Lines of descent are not broken, because the population range expansion events were accompanied by statistically significant admixture when they involved expansion into a previously inhabited area. Significant inferences about human evolution from multilocus, nested-clade phylogeographic analysis. Geographic location is indicated on the x-axis, and time on the y-axis, with the bottom of the figure corresponding to 2 million years ago. Vertical lines indicate genetic descent over time, and diagonal lines indicate gene flow across space and time. Thick arrows indicate statistically significant further genetic interchange between African and Eurasian populations, further weakening the hypothesis of isolated Pleistocene lineages of humans. Gene flow then continued until a third major expansion of humans out of Africa into Eurasia occurred around 130,000 years ago, the time of the last major Saharan wet period. The fossil record indicates that modern humans began expanding out of sub-Saharan Africa at 130,000 years ago and reached China no later than 110,000 years ago. The null hypothesis of no admixture is overwhelmingly rejected for this expansion event. Following the expansion with admixture of modern humans from Africa, there have been additional expansions, mostly into areas not formerly occupied by humans (figure 2). Wherever humans lived, gene flow was established, mostly limited by isolation by distance but with some long-distance dispersal as well. On a timescale of tens of thousands of years, there is not one statistically significant inference of splitting or isolation during the last 700,000 years. Sometimes, environmental factors vary over the geographic range of the species, and some of these environmental factors can induce natural selection that results in local adaptation. This reasoning leads to the idea that local adaptations can sometimes be biological markers of racial status. Variation in environmental factors can still induce natural selection that results in local adaptations in species with sufficient gene flow and admixture to prevent race formation. However, in this case, the geographic distributions of the local adaptations reflect the geography of the environmental factors and not racial boundaries. Frequently, different adaptive traits display discordant geographic distributions, thereby indicating that these are simply adaptations of local populations and not markers of higher groupings such as race. Because humans are not subdivided into races by any of the definitions applied to other species, the locally adaptive traits of humans are not "racial" traits. Skin color is historically the locally adaptive trait most commonly considered a "racial" trait in humans. Skin color varies continuously among humans and does not fall into a few discrete "racial" types. For example, the native peoples with the darkest skins live in tropical Africa and Melanesia. Because Africans and Melanesians live on opposite sides of the world, they are more highly genetically differentiated than many other human populations (figure 1). Hence, skin color is not an indicator of the degree of genetic differentiation, as a true racial trait would be. Another adaptive trait found in Africa is resistance to sleeping sickness, and the responsible gene is found at frequencies up to 80 percent in parts of western Africa where the parasite that causes sleeping sickness is common. Hence, the distribution of sleeping sickness resistance is only a subset of the geographic distribution of dark skin in Africa. Another adaptive trait is resistance to malaria, which is widespread in African populations. However, malaria is also common in some areas outside Africa, and malarial resistance is found in many European and Asian populations as well. Indeed, one of the alleles underlying malarial resistance, the sickle-cell allele, is most frequent in certain populations on the Arabian Peninsula despite often being regarded as a disease of "blacks. The discordance in the distributions of adaptive traits in humans makes them useless in defining races. Human evolutionary history has been dominated by gene flow and admixture that unifies humanity into a single evolutionary lineage. Isolation by distance ensures that human populations are genetically differentiated from one another, and local adaptation ensures that some of these differences reflect adaptive evolution to the environmental heterogeneity that our globally distributed species experiences. However, most of our genetic variation exists as differences among individuals, with between-population differences being very small. There are no biological races in humans; indeed, despite our global distribution, we are one of the most genetically homogeneous species on this planet. Evidence from Cameroon reveals differences in the genetic structure and histories of chimpanzee populations. Human races in the context of recent human evolution: A molecular genetic perspective. This paper covers many of the same issues as this chapter, but with older data sets. The results of multilocus nested clade phylogeographic analysis of humans, showing that gene flow and admixture have been such common features of recent human evolution that there is only one evolutionary lineage of humanity. The authors strongly argue against typological thinking in anthropology that explains human variation in terms of a few types or "races" rather than dealing with the entire range of variation found in living humans. Although the book ignores the nonhuman literature and tends to hyperbole, it remains an excellent introduction to the intertwined topics of racism and models of modern human origins. Formal statistics tests of the hypothesis that human genetic variation is structured in a treelike fashion. They show that the hypothesis of a tree of populations is strongly rejected for humans. Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa Relethford, J. This paper shows that skin color variation and other morphological traits are clinal and are not well described by discrete racial categories. Future nonadaptive evolution Future adaptive evolution Eugenics and genetic engineering How humans will evolve in the future is highly speculative because the process of evolution depends critically on random processes such as mutation, recombination, and genetic drift, and because adaptive evolution is strongly influenced by changing environments. Because the human environment includes culture, which can change quickly, it is difficult to predict future environments and hence future adaptive evolution. Nevertheless, some predictions can be made based on a basic understanding of evolutionary mechanisms. An allele that is functionally equivalent to its ancestral allele in terms of its chances of being replicated and passed on to the next generation. The evolutionary force associated with random sampling events that alters the frequencies of genetic variants in the gene pool. The deliberate modification of characteristics of an organism by manipulating its genetic material. The condition in which the two homologous segments of genetic material inherited from the parents are of a different state. We can infer to some extent what our ancestors were like 100,000 or a million years ago, but what will our descendants be like 100,000 or a million years into the future? This question is difficult to answer because the evolutionary process itself is strongly influenced by random factors. There can be no evolution of any sort without genetic variation, and this genetic variation is created by mutation and recombination.
4 mg cyproheptadine mastercard. Dry Cough: Symptoms and Treatment.